|

Syndrome
ImmunoDéficitaire Acquis (SIDA)
3.
Le virus
Luc
Montagnier/Institut Pasteur/CNRI/Science Source/Photo
Researchers, Inc.
Virus
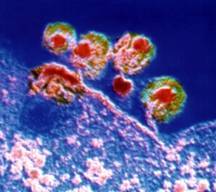
du sida
Particules virales du VIH
vues en microscopie électronique et en fausses couleurs
à la surface d'une cellule infectée.
Encyclopédie Microsoft ® Encarta ® 2002. © 1993-2001
Microsoft Corporation. Tous droits réservés.
Le HIV-1, virus responsable
du sida, a été découvert en 1983 par une équipe
de virologistes de l'Institut Pasteur (Paris),
dirigée par le professeur Luc Montagnier. Robert
Gallo et Max Essex, chercheurs au National Cancer
Institute de Bethesda (Maryland), ont découvert
simultanément l'agent causal de la maladie, qu'ils
ont baptisé HTLV-III. Les deux virus se sont révélés
identiques. Une querelle juridique a opposé les
deux instituts de recherche afin d'établir l'antériorité
d'une équipe par rapport à l'autre et les droits
financiers sur les tests de diagnostic mis en
place pour le dépistage de la maladie. Il est
aujourd'hui établi que la légitimité de la découverte
revient à la France. Les deux instituts ont signé
un accord à l'amiable en 1987, mettant un terme
au conflit. Toutefois, Robert Gallo et son équipe
ont mis en évidence le TCGF (T cell growth
factor), appelé interleukine 2, qui a
permis la mise en culture des lymphocytes T
et a conduit à la découverte en 1978 du premier
rétrovirus humain, le HTLV-1 (Human T
lymphotropic virus 1), et en 1982 à celle
du HTLV-2.
3.1.
Les particularités du virus
©
Microsoft Corporation. Tous droits réservés.
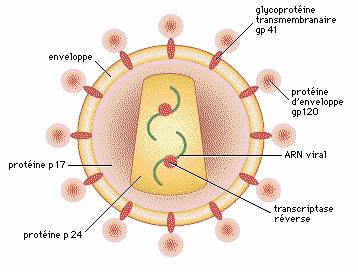
Structure du virus du sida
Le virus du sida (HIV) présente, à sa surface, des glycoprotéines
d'enveloppe (gp 120) qui lui permettent de se fixer
spécifiquement sur les lymphocytes T-CD4 et des
macrophages, par l'intermédiaire des molécules CD4
que portent ces derniers. Les glycoprotéines gp 40
sont impliquées dans la fusion de l'enveloppe du virus
avec la membrane de ces cellules. Une enzyme, la transcriptase
réverse, permet la transcription du matériel génétique
du HIV, formé d'ARN, en ADN, lequel sera ensuite intégré
au génome cellulaire. Les autres protéines du virus
sont des protéines de structure.
Encyclopédie Microsoft ® Encarta ® 2002. © 1993-2001
Microsoft Corporation. Tous droits réservés.
Classé dans le groupe
des rétrovirus et le sous-groupe des lentivirus, le
virus du sida possède un matériel génétique constitué
d'ARN. La structure de son patrimoine génétique est
différente de celle de la majorité des rétrovirus. En
effet, en plus des gènes de structure communs aux autres
rétrovirus (gag, pol et env), il possède 6 autres
gènes (tat, rev, nef, pvu, vpr et vif), dont les produits
ont un rôle régulateur. Le gène gag contient l'information
nécessaire à la synthèse de glycoprotéines du virus.
Le gène pol code pour des protéines nécessaires à la
« rétrotranscription » du matériel génétique
viral ; il code en particulier pour une enzyme,
la transcriptase reverse, capable de recopier l'ARN
en ADN, phénomène indispensable à sa multiplication
et à son intégration dans le génome des cellules. Le
gène env code pour les protéines de l'enveloppe virale.
L'équipe française du Dr Klatzman a démontré que
le HIV a pour cible les lymphocytes CD4. Le virus
peut infecter les macrophages, globules blancs qui alertent
les T4, en cas de présence d'un corps étranger, en les
utilisant comme réservoir.
3.2.
Multiplication du virus
CNRI/Science Source/Photo Researchers,
Inc.
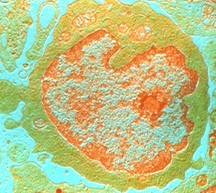
Lymphocyte
Produits par la moelle osseuse, les lymphocytes sont
des cellules appartenant au système immunitaire. Les
lymphocytes B produisent les protéines de défense,
les anticorps, qui se fixent aux substances étrangères
à l'organisme (antigènes), permettant leur élimination.
Outre cette forme d'immunité dite humorale, l'immunité
cellulaire met en jeu d'autres lymphocytes, les lymphocytes T,
dans une série de réactions d'activation du système
immunitaire en cas d'infection. C'est le cas notamment
des lymphocytes T auxiliaires, que le virus du
sida détruit en premier lieu.
Encyclopédie Microsoft ® Encarta ® 2002. © 1993-2001
Microsoft Corporation. Tous droits réservés.
Comme tous les virus,
le HIV est un parasite : il ne se reproduit qu'au
sein d'une cellule hôte qu'il détruit. Le virus s'attache
à la cellule cible (lymphocyte CD4) puis pénètre
dans le cytoplasme. L'ARN viral, matériel génétique
du HIV, est transcrit en ADN proviral grâce à l'enzyme
spécifique : la transcriptase inverse. L'ADN proviral
atteint le noyau de la cellule colonisée où il s'intègre
à ses chromosomes et produit des milliers de copies
d'ARN viral, dont les ARN messagers synthétisant les
protéines virales qui, après un assemblage méticuleux,
constitueront le virus complet. Ce dernier acquiert
ses facultés infectieuses grâce à l'action d'une enzyme,
la protéase, qui l'active en agissant sur une protéine
de la coque virale. Parmi les virus produits par une
cellule hôte, plusieurs sont imparfaits et par conséquent
non infectieux. Pour se multiplier, on dit que le virus
se réplique (réplication du matériel génétique). L'infection
à HIV est définitive et persistante, car la production
de particules virales en plus ou moins grande quantité
est continue.
3.3.
Diversité du virus
En l'état actuel des connaissances,
on note une variabilité du HIV. Dès 1985, on a pu identifier
deux types de virus, le HIV-1 et le HIV-2.
Leur différence réside au niveau des séquences géniques.
Le HIV-1 est le plus répandu dans le monde. L'analyse
génétique distingue dans le HIV-1 deux groupes :
le groupe M (pour majeur) et le groupe O (pour
ouiller). Le groupe M du HIV-1 est divisé en huit
sous-groupes de A à H. Comme il existe plusieurs configurations
possibles de recombinaison, un individu peut être infecté
par deux ou plusieurs souches à la fois. Le groupe O
du HIV-1 a été isolé chez des patients originaires de
certaines régions d'Afrique centrale.
La situation thaïlandaise
évoque deux épidémies parallèles, la première due au
sous-type E, correspondant à une transmission par
voie sexuelle, et la seconde due au sous-groupe B,
correspondant à une transmission sanguine liée à une
toxicomanie avec injection intraveineuse. Le HIV-2 peut
également être typé en sous-groupes. On note qu'il est
limité à l'Afrique de l'Ouest et qu'une proximité géographique
avec les forêts tropicales et l'habitat de singes infectés
par le lentivirus SIV (virus proche du HIV-2) conduit
à se poser un certain nombre de questions sur l'origine
épidémiologique de la maladie (interespèces). La variation
antigénique soulève des points importants quant au typage
des virus et à l'évaluation de la fréquence de recombinaison.
Source:
Encyclopédie Microsoft ® Encarta ® 2002. © 1993-2001
Microsoft Corporation. Tous droits réservés.
|